恒温动物包括人,有完善的体温调节机制。在外界环境温度改变时,通过调节产热过程和散热过程,维持体温相对稳定。例如,在寒冷环境下,机体增中产热和减少散热;在炎热环境下,机体减少产热和增加散热,从而使体温保持相对稳定。这是复杂的调节过程,涉及感受温度变化的温度感觉器,通过有关传导通路把温度信息传达到体温调节中枢,经过中枢整合后,通过自主神经系统调节皮肤血流量、竖毛肌和汗腺活动等;通过躯体神经调节骨骼肌的活动,如寒战等;通过内分泌系统,改变机体的代谢率。
体温调节是生物自动控制系统的实例。如图7-8所示,下丘脑体温调节中枢,包括调定点(set point)神经元在内,属于控制系统。它的传出信息控制着产热器官如肝、骨骼肌以及散热器官如皮肤血管、汗腺等受控系统的活动,使受控对象――机体深部温度维持一个稳定水平。而输出变量体温总是会受到内、旬环境因素干扰的(譬如机体的运动或外环境气候因素的变化,如气温、湿度、风速等)。此时则通过温度检测器――皮肤及深部温度感受器(包括中枢温度感受器)将干扰信息反馈于调定点,经过体温调节中枢的整合,再调整受控系统的活动,仍可建立起当时条件下的体热平衡,收到稳定体温的效果。
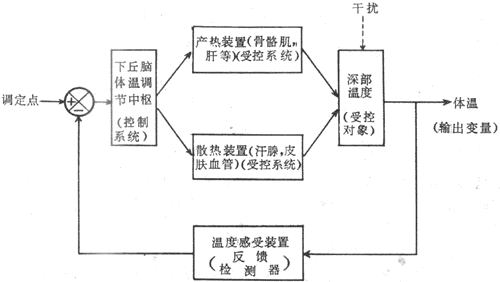
图7-8 体温调节自动控制示意图
(一)温度感受器对温度敏感的感受器称为温度感受器,温度感受器分为外周温度感受器和中枢温度感受器。
外周温度感受器在人体皮肤、粘膜和内脏中,温度感受器分为冷感受器和温觉感受器,它们都是游离神经末稍的。当皮肤温度升高时,温觉感受器兴奋,而当皮肤温度下降时,则冷感受器兴奋。从记录温度感受器发放冲动可看到,温觉感受器和冷觉感受器在28℃时发放冲动频率最高,而温觉感受器则在43℃时发放冲动频率最高。当皮肤温度偏离这两个温度时,两种感受器发放冲动的频率都逐渐下降(图7-9)。此外,温度感受器对皮肤温度变化速率更敏感。
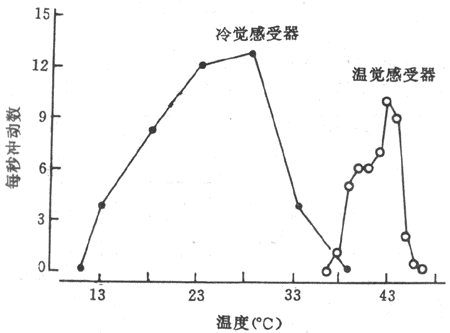
图7-9 大鼠阴囊皮肤冷觉和温觉感受器
内脏器官也有温度感受器。有人将电热器埋藏在差点羊腹腔内 并加温至43-44℃,观察到羊的呼吸频率和蒸发散热迅速增加,加热3-5分钟后,动物开始喘息,使下丘脑温度下降。说明内脏温度升高可引起明显的散热反应。
中枢温度感受器在脊髓、延髓、脑干网状结构及下丘脑中有温度感受器。
用改变脑组织温度的装置(变温管),对不麻醉或麻醉的兔、猫或狗等的下丘脑前部进行加温或冷却,发现在视前区-下丘脑前部(preoptic anterior hypothalamus,PO/AH)加温,可引起动物出现喘息和出汗等散热反应,而局部冷却则引起产热量增加,说明PO/AH本身就可调节散热和产热这两种相反的过程。用电生理方法记录PO/AH中存在着热敏神经元(warm-sensitive neuron)和冷敏神经元(cold-sensitiveneuron)。前者的放电频率随局部温度的升高而增加,而后者的放电频率则随着脑组织的降温而增加(图7-10,7-11)。实验证明,局部脑组织温度变动0.1C,这两种温度敏感神经元的放电频率就会反映出来,而且不出现适应现象。
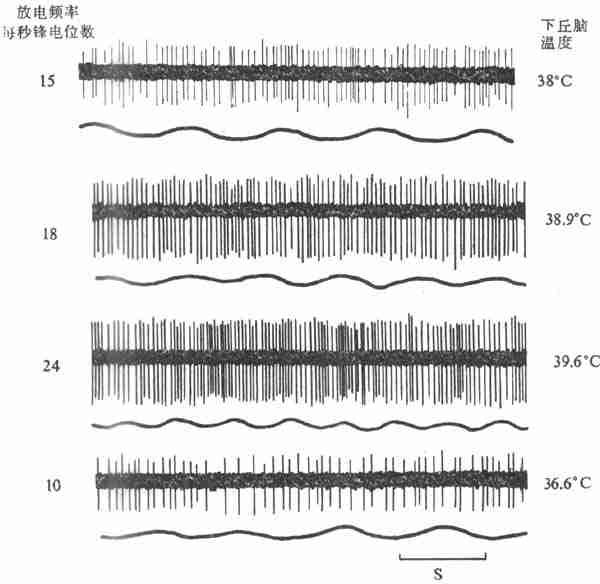
图7-10下丘脑局部加温时热敏神经元放电的记录(上)和呼吸曲线(下)(猫)
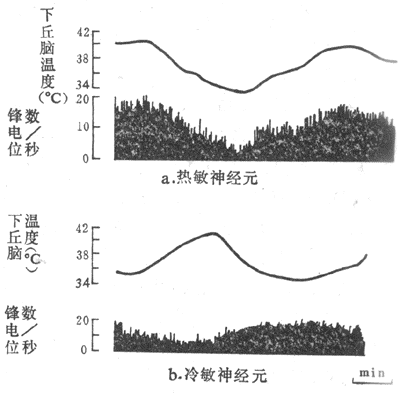
图7-11由下丘脑视前区导出的温度敏感神经元的放电活动
脊髓中也有温度敏感神经元。冷却轻度麻醉狗的颈、胸髓或胸腰髓,则动物出现皮肤血管收缩和寒战等体温调节反应。这时,切断被冷却部位的后根或高位切断脊髓,血管反应和寒战也不消失。加温脊髓,则引起皮肤血管舒张和热喘呼吸,寒战受到抑制。另外,据谓脊髓中传导温度信息的上行性神经元的纤维前侧侧索中走行,它将信息发送给PO/AH。
延髓中也存在着温度敏感神经元。皮肤、脊髓及中脑的传入温度信息都会聚于延髓温度敏感神经元;而延髓也接受来自PO/AH的信息,并且向PO/AH输送信息。
脑干网状结构也有对局部温度变化发生反应的神经元,它接受发生皮肤、脊髓的温度信息,并且向PO/AH输送温度信息。
(二)体温调节中枢根据多种恒温动物脑的分段切除实验看到,切除大脑皮层及部分皮层下结构后,只要保持下丘脑及其以下的神经结构完整,动物虽然在行为方面可能出现一些欠缺,但仍具有维持恒定体温的能力。如进一步破坏下丘脑,则动物不再具有维持体温相对恒定的能力。这些事实说明,调节体温的基本中枢在下丘脑。下丘脑局部破坏或电刺激等实验观察到,PO/AH破坏,则散热反应消失,体温升高;刺激之,则引起散热反应,而且寒战受到抑制;而破坏下丘脑后部,体温下降,产热反庆受抑制;刺激之,则引起寒战。据此得出结论,下丘脑前部是散热中枢,而下丘脑后部是产热中枢,但是,这两种实验方法比较粗糙,因此得出来的结论也较精细的实验方法观察到的结果不相符。
前已述及, PO/AH就有热敏神经元和冷敏神经元,分别调节散热和产热反应。下丘脑以外的脑细胞也有类似的两种神经元存在。看来没有明确定位的产热中枢和散热中枢。体温调节是涉及多方输入温度信息和多系统的传出反应,因此是一种高级的中枢整合作用。视前区-下丘脑前部应是体温调节的基本部位。下丘脑前部的热敏神经元和冷敏神经元既能感受它们所在部位的温度变化,又能过传入的温度信息进行整合。因此,当外界环境温度改变时,可通过①皮肤的温、冷觉感受器的刺激,将温度变化的信息沿躯体传入神经经脊髓到达下丘脑的体温调节中枢;②外界温度改变可通过血液引起深部温度改变,并直接作用于下丘脑前部;③脊髓和下丘脑以外的中枢温度感受器也将温度信息传给下丘脑前部。通过下丘脑前部和中枢其它部位的整合作用,由下述三条途径发出指令调节体温:①通过交感神经系统调节皮肤血管舒缩反应和汗腺分泌;②通过躯体神经改变骨骼肌的活动,如在寒冷环境时的寒战等;③通过甲状腺和肾上腺髓质的激素分泌活动的改变来调节机体的代谢率。有人认为,皮肤温度感受器兴奋主要调节皮肤血管舒活动和血流量;而深部温度改变则主要调节发汗和骨骼肌的活动。通过上述的复杂调节过程,使机体在外界温度改变时能维持体温相对稳定。
调定点学说 此学说认为,体温的调节类似于恒温器的调节,PO/AH中有个调定点,即规定数值(如37℃)。如果偏离此规定数值,则由反馈系统将偏离信息输送到控制系统,然后经过对受控系统的调整来维持体温的恒定。通常认为,PO/AH中的温度敏感神经元可能在体温调节中起着调定点的作用。例如,此学说认为,由细菌所致的发热是由于热敏神经元的阈值因受到热原(pyrogen)的作用而升高,调定点上移(如39℃)的结果。因此,发热反应开始先出现恶寒战栗等产热反应,直到体温升高到39℃以上时才出现散热反应。只要致热因素不消除,产热与散热两个过程就继续在此新的体温水平上保持着平衡。应该指出的是,发热时体温调节功能并无阻碍,而只是由于调定点上移,体温才被调节到发热水平。
单胺物质对体温调节的作用在哺乳动物下丘脑的与体温调节有关的神经末稍中含有丰富的单胺物质。60年代初,用狗、猫、猴做的实验证明,用5-羧色胺灌注动物的脑室或微量注入于下丘脑,动物的体温上升,同时伴有血管收缩反应和寒战;而去甲肾上肾素则使动物的体温降低0.5-2℃,同时伴有外周血管舒张。根据这类实验,提出了体温调节的单胺学说,此学说认为,5-羧色胺和去甲肾上腺素这两种物质在量上的动态平衡可保持体温的恒定。但目前认为,这两种物质对体温调节中枢的活动只能起到调整的作用,而对于体温的恒定水平没有决定作用。